
 The paper is behind a paywall, but there is plentiful raw genetic data in the online supplement. I'll probably have much more to say on this when I read it, but here's the groundbreaking part:
The paper is behind a paywall, but there is plentiful raw genetic data in the online supplement. I'll probably have much more to say on this when I read it, but here's the groundbreaking part:Most of this sample belonged to haplogroup G2a-P15 with some I2a-P37.2 also represented.
G2a was also one of the haplogroups represented in a small sample from Neolithic Central Europe. I think we can now safely say that G2a may have been the main Neolithic link that ties the farmers that went north across the Balkans to Central Europe, and those that followed the western, maritime route to the Western Mediterranean. The unambiguous West Asian origin of this lineage should put to rest any ideas about Neolithic farmers in the Western Mediterranean being descended from indigenous Mesolithic foragers.
I-P37.2 is also quite interesting, as it is tied to the Balkans, but also modern Southwestern Europe (it is especially frequent in Sardinia in its derived M26+ form). ISOGG tells me that:
I2-M438 et al includes I2* which shows some membership from Armenia, Georgia and Turkey; I2a-P37.2, which is the most common form in the Balkans and Sardinia. I2a1-M26 is especially prevalent in Sardinia. I2b-M436 et al reaches its highest frequency along the northwest coast of continental Europe. I2b1-M223 et al occurs in Britain and northwest continental Europe. I2b1a-M284 occurs almost exclusively in Britain, so it apparently originated there and has probably been present for thousands of years.
If these aren't signals of a maritime pioneer colonization that followed the maritime route along the Mediterranean and Atlantic, I don't know what is.

 The Y-chromosome is an extreme outlier compared to modern groups, probably because of its heavy G2a domination, whereas the mtDNA from Treilles appears just like a normal and unexceptional Mediterranean-type population.
The Y-chromosome is an extreme outlier compared to modern groups, probably because of its heavy G2a domination, whereas the mtDNA from Treilles appears just like a normal and unexceptional Mediterranean-type population.
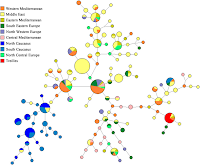
UPDATE V: The G2a median joining network shows that the Treilles haplotypes are disjoint from those that dominate the North Caucasus, with clear links to the Middle East, Central/East Mediterranean regions, as well as the South Caucasus.
Ancient DNA reveals male diffusion through the Neolithic Mediterranean route
Marie Lacan et al.
The Neolithic is a key period in the history of the European settlement. Although archaeological and present-day genetic data suggest several hypotheses regarding the human migration patterns at this period, validation of these hypotheses with the use of ancient genetic data has been limited. In this context, we studied DNA extracted from 53 individuals buried in a necropolis used by a French local community 5,000 y ago. The relatively good DNA preservation of the samples allowed us to obtain autosomal, Y-chromosomal, and/or mtDNA data for 29 of the 53 samples studied. From these datasets, we established close parental relationships within the necropolis and determined maternal and paternal lineages as well as the absence of an allele associated with lactase persistence, probably carried by Neolithic cultures of central Europe. Our study provides an integrative view of the genetic past in southern France at the end of the Neolithic period. Furthermore, the Y-haplotype lineages characterized and the study of their current repartition in European populations confirm a greater influence of the Mediterranean than the Central European route in the peopling of southern Europe during the Neolithic transition.
Link
What is absent is also quite interesting as what is present. The absence of E1b1b is consistent with my theory about the Bronze Age Greek expansion of that haplogroup in Europe that has been tied to the historical Greeks of the West Mediterranean.
R-M269 which, because of its apparent young Y-STR age has been tied by some to either the Mediterranean or Central European Neolithic is conspicuous absently from both at the moment. It may yet surface in a Neolithic context, but its absence this late from a region where, today, it is abundant only adds to its mystery. The absence of J2 is equally mysterious, as this is another putative Neolithic lineage which has failed to appear so far in a Neolithic context, while its J1 sister clade did make an appearance in much later aboriginals from the Canary Islands.
UPDATE I: Interestingly, some French researchers had noted a littoral distribution of haplogroups I, J, G in the Finistère, on the Atlantic side.
UPDATE II: I was reviewing my Ancient Y-chromosome studies compendium and one thing starts to become clear: how many of the earliest samples we have were dominated by 1-2 haplogroups, whereas there is a plethora of haplogroups in most modern populations: Treilles, Krasnoyarsk, Xiaohe, Pengyang all belonged to a single haplogroup, while Yangtze China to several lineages, all of which were in the O haplogroup.
Look at the MDS plot of the Y-chromosome and mtDNA from Treilles:

 The Y-chromosome is an extreme outlier compared to modern groups, probably because of its heavy G2a domination, whereas the mtDNA from Treilles appears just like a normal and unexceptional Mediterranean-type population.
The Y-chromosome is an extreme outlier compared to modern groups, probably because of its heavy G2a domination, whereas the mtDNA from Treilles appears just like a normal and unexceptional Mediterranean-type population.Perhaps the modern Caucasus where particular ethnic groups are dominated by particular Y-haplogroups is a good analogy for prehistoric man, with many different groups with their signature haplogroups kept disjoint patrilineal gene pools before beginning to merge in late prehistorical and historical times.
UPDATE III: A poster at dna-forums as well as Ken Nordtvedt both agree that the I2a haplotypes belong to I-M26, a haplogroup that is modal in the SW Mediterranean, reaching very high frequencies in Sardinia. This may be consistent with the great biological continuity since the Neolithic in Sardinia, continuity which is also evident on the mtDNA. It also shows why the inference of pre-Neolithic I2a in Sardinia was flawed because of the use of the evolutionary mutation rate, while the origin and expansion of I-M26 in "genealogical rate" years becomes 5-7ky, consistent with the Neolithic origin of that haplogroup and the ancient DNA presence in Neolithic France.
UPDATE IV: Table S4 lists (in %) shared mtDNA lineages between Treilles and modern populations. The top ones are: Welsh (17.391), Cornish (16.667), Central Greeks (14.286), Bulgarians (12.5). Several Italian groups as well as South Tyrol Ladins and Germans are also greater than 10%.

UPDATE V: The G2a median joining network shows that the Treilles haplotypes are disjoint from those that dominate the North Caucasus, with clear links to the Middle East, Central/East Mediterranean regions, as well as the South Caucasus.
UPDATE VI: Some more good news: "The ancient DNA Lacan is now extracting from skeletons across France and Spain, Haak says, should provide more “piece[s] of the enormous puzzle we are trying to put together.”
PNAS doi: 10.1073/pnas.1100723108
Ancient DNA reveals male diffusion through the Neolithic Mediterranean route
Marie Lacan et al.
The Neolithic is a key period in the history of the European settlement. Although archaeological and present-day genetic data suggest several hypotheses regarding the human migration patterns at this period, validation of these hypotheses with the use of ancient genetic data has been limited. In this context, we studied DNA extracted from 53 individuals buried in a necropolis used by a French local community 5,000 y ago. The relatively good DNA preservation of the samples allowed us to obtain autosomal, Y-chromosomal, and/or mtDNA data for 29 of the 53 samples studied. From these datasets, we established close parental relationships within the necropolis and determined maternal and paternal lineages as well as the absence of an allele associated with lactase persistence, probably carried by Neolithic cultures of central Europe. Our study provides an integrative view of the genetic past in southern France at the end of the Neolithic period. Furthermore, the Y-haplotype lineages characterized and the study of their current repartition in European populations confirm a greater influence of the Mediterranean than the Central European route in the peopling of southern Europe during the Neolithic transition.
Link

















