This seems to be in wonderful agreement with the coalescence ages of haplogroups M1 and U6. I would say that some type of Into-Africa migration makes the best sense of the totality of the evidence, now that we know that the Iberomaurusian appears after a hiatus in occupation.
Journal of Human Evolution doi:10.1016/j.jhevol.2013.06.003
Origins of the Iberomaurusian in NW Africa: New AMS radiocarbon dating of the Middle and Later Stone Age deposits at Taforalt Cave, Morocco
R.N.E. Barton et al.
Recent genetic studies based on the distribution of mtDNA of haplogroup U6 have led to subtly different theories regarding the arrival of modern human populations in North Africa. One proposes that groups of the proto-U6 lineage spread from the Near East to North Africa around 40–45 ka (thousands of years ago), followed by some degree of regional continuity. Another envisages a westward human migration from the Near East, followed by further demographic expansion at ∼22 ka centred on the Maghreb and associated with a microlithic bladelet culture known as the Iberomaurusian. In evaluating these theories, we report on the results of new work on the Middle (MSA) and Later Stone (LSA) Age deposits at Taforalt Cave in Morocco. We present 54 AMS radiocarbon dates on bone and charcoals from a sequence of late MSA and LSA occupation levels of the cave. Using Bayesian modelling we show that an MSA non-Levallois flake industry was present until ∼24.5 ka Cal BP (calibrated years before present), followed by a gap in occupation and the subsequent appearance of an LSA Iberomaurusian industry from at least 21,160 Cal BP. The new dating offers fresh light on theories of continuity versus replacement of populations as presented by the genetic evidence. We examine the implications of these data for interpreting the first appearance of the LSA in the Maghreb and providing comparisons with other dated early blade and bladelet industries in North Africa.
Link

July 31, 2013
West Eurasian admixture in Khoe-San via East Africa
A new paper on the arXiv quantifies and dates the West Eurasian admixture in east Africa, and uncovers the presence of such admixture even in the Khoe-San of southern Africa. It appears that the admixture first occurred in East Africa about ~3ky ago, and reached southern Africa about ~1.5ky ago.

It is quite remarkable that different waves of migration converged into southern Africa from different directions: west African farmers and west Eurasian-admixed east African pastoralists. We should count ourselves lucky that the Khoe-San were discovered when they did: a few centuries more, and they too might have followed the fate of other populations finding themselves at the losing side of a technology differential, their culture lost, and their DNA preserved only as fragments in the gene pools of the more successful groups.
arXiv:1307.8014 [q-bio.PE]
Ancient west Eurasian ancestry in southern and eastern Africa
Joseph K. Pickrell et al.
The history of southern Africa involved interactions between indigenous hunter-gatherers and a range of populations that moved into the region. Here we use genome-wide genetic data to show that there are at least two admixture events in the history of Khoisan populations (southern African hunter-gatherers and pastoralists who speak non-Bantu languages with click consonants). One involved populations related to Niger-Congo-speaking African populations, and the other introduced ancestry most closely related to west Eurasian (European or Middle Eastern) populations. We date this latter admixture event to approximately 900-1,800 years ago, and show that it had the largest demographic impact in Khoisan populations that speak Khoe-Kwadi languages. A similar signal of west Eurasian ancestry is present throughout eastern Africa. In particular, we also find evidence for two admixture events in the history of Kenyan, Tanzanian, and Ethiopian populations, the earlier of which involved populations related to west Eurasians and which we date to approximately 2,700 - 3,300 years ago. We reconstruct the allele frequencies of the putative west Eurasian population in eastern Africa, and show that this population is a good proxy for the west Eurasian ancestry in southern Africa. The most parsimonious explanation for these findings is that west Eurasian ancestry entered southern Africa indirectly through eastern Africa.
Link

It is quite remarkable that different waves of migration converged into southern Africa from different directions: west African farmers and west Eurasian-admixed east African pastoralists. We should count ourselves lucky that the Khoe-San were discovered when they did: a few centuries more, and they too might have followed the fate of other populations finding themselves at the losing side of a technology differential, their culture lost, and their DNA preserved only as fragments in the gene pools of the more successful groups.
arXiv:1307.8014 [q-bio.PE]
Ancient west Eurasian ancestry in southern and eastern Africa
Joseph K. Pickrell et al.
The history of southern Africa involved interactions between indigenous hunter-gatherers and a range of populations that moved into the region. Here we use genome-wide genetic data to show that there are at least two admixture events in the history of Khoisan populations (southern African hunter-gatherers and pastoralists who speak non-Bantu languages with click consonants). One involved populations related to Niger-Congo-speaking African populations, and the other introduced ancestry most closely related to west Eurasian (European or Middle Eastern) populations. We date this latter admixture event to approximately 900-1,800 years ago, and show that it had the largest demographic impact in Khoisan populations that speak Khoe-Kwadi languages. A similar signal of west Eurasian ancestry is present throughout eastern Africa. In particular, we also find evidence for two admixture events in the history of Kenyan, Tanzanian, and Ethiopian populations, the earlier of which involved populations related to west Eurasians and which we date to approximately 2,700 - 3,300 years ago. We reconstruct the allele frequencies of the putative west Eurasian population in eastern Africa, and show that this population is a good proxy for the west Eurasian ancestry in southern Africa. The most parsimonious explanation for these findings is that west Eurasian ancestry entered southern Africa indirectly through eastern Africa.
Link
July 25, 2013
Middle Paleolithic and Neolithic occupations around Mundafan palaeolake (Crassard et al. 2013)

Survey in the Mundafan palaeolake basin revealed, for the first time, Middle Palaeolithic occupations. Recovery of Middle Palaeolithic assemblages corresponds with recent environmental and geoarchaeological studies that indicate at least three lacustrine wet phases in MIS 5 [23]. The main diagnostic lithic technology observed is the preferential Levallois reduction method, which is also present at the Jubbah palaeolake during MIS 5 (JQ-1, JSM-1 and JKF-1 sites: [21], [22]) and at the Jebel Faya rock shelter at the transition between MIS 6–5e (Assemblage C: [6]). This technology is absent in MIS 3 in southwest Yemen (SD1, SD2 and AS1 sites: [8], [9]). Earlier dating for preferential Levallois in Arabia, in MIS 7, is possible, but still insufficiently represented owing to small sample size at the Jubbah palaeolake (JQ-1: [22]). Other preferential Levallois methods have been observed in Dhofar, including in the Nubian Complex, dated to at least ca. 106 ka [7], in Hadramawt and the southern fringe of the Rub’ al-Khali, Oman [10], and in central Saudi Arabia at Al-Kharj [75]. Nubian Complex technology has not yet been identified at Mundafan. We associate the Levallois component in Mundafan with the wet pluvials of MIS 5, most probably during the wetter events of MIS 5e (ca. 125 ka), MIS 5c (ca. 100 ka) and MIS 5a (80 ka), when conditions were more favorable for hominin dispersals. The Middle Palaeolithic evidence thus provides empirical support for Rosenberg and colleagues assertion [23] that the dispersal of hominins into the Rub’ al-Khali occurred during ameliorated periods, and perhaps supports their claim for the expansion of Homo sapiens into this marginal environment.PLoS ONE 8(7): e69665. doi:10.1371/journal.pone.0069665
Middle Palaeolithic and Neolithic Occupations around Mundafan Palaeolake, Saudi Arabia: Implications for Climate Change and Human Dispersals
Rémy Crassard et al.
The Arabian Peninsula is a key region for understanding climate change and human occupation history in a marginal environment. The Mundafan palaeolake is situated in southern Saudi Arabia, in the Rub’ al-Khali (the ‘Empty Quarter’), the world’s largest sand desert. Here we report the first discoveries of Middle Palaeolithic and Neolithic archaeological sites in association with the palaeolake. We associate the human occupations with new geochronological data, and suggest the archaeological sites date to the wet periods of Marine Isotope Stage 5 and the Early Holocene. The archaeological sites indicate that humans repeatedly penetrated the ameliorated environments of the Rub’ al-Khali. The sites probably represent short-term occupations, with the Neolithic sites focused on hunting, as indicated by points and weaponry. Middle Palaeolithic assemblages at Mundafan support a lacustrine adaptive focus in Arabia. Provenancing of obsidian artifacts indicates that Neolithic groups at Mundafan had a wide wandering range, with transport of artifacts from distant sources.
Link
Nubian complex site from central Arabia (Crassard & Hilbert 2013)

The palaeoclimatic record of Arabia indicates that three distinct wet phases occurred during MIS 5 [109]. The first of these wet phases occurred between 130 and 125 kya (MIS 5e) and precedes the presence of Nubian technology in Arabia. The two following wet phases, positioned around 100 kya (MIS 5c) and between 80 to 75 kya (MIS 5a) may be viewed as possible windows for the Nubian expansion into and across Arabia (Figure 11).PLoS ONE 8(7): e69221. doi:10.1371/journal.pone.0069221
A Nubian Complex Site from Central Arabia: Implications for Levallois Taxonomy and Human Dispersals during the Upper Pleistocene
Rémy Crassard, Yamandú Hieronymus Hilbert
Archaeological survey undertaken in central Saudi Arabia has revealed 29 surface sites attributed to the Arabian Middle Paleolithic based on the presence of Levallois blank production methods. Technological analyses on cores retrieved from Al-Kharj 22 have revealed specific reduction modalities used to produce flakes with predetermined shapes. The identified modalities, which are anchored within the greater Levallois concept of core convexity preparation and exploitation, correspond with those utilized during the Middle Stone Age Nubian Complex of northeast Africa and southern Arabia. The discovery of Nubian technology at the Al-Kharj 22 site represents the first appearance of this blank production method in central Arabia. Here we demonstrate how a rigorous use of technological and taxonomic analysis may enable intra-regional comparisons across the Arabian Peninsula. The discovery of Al-Kharj 22 increases the complexity of the Arabian Middle Paleolithic archaeological record and suggests new dynamics of population movements between the southern and central regions of the Peninsula. This study also addresses the dichotomy within Nubian core typology (Types 1 and 2), which was originally defined for African assemblages.
Link
July 22, 2013
New A00 project
A new project dedicated to Y-haplogroup A00 is seeking funding. A00 is the most basal clade of the human Y-chromosome phylogeny and has so far been found in African Americans and Mbo people from Cameroon. This is how the funds raised will be used:
In this campaign, we're seeking the funds we need to launch our first phase of fieldwork in late August, 2013. Our overall research plan includes five field trips to sample peoples in different regions of the country. In the first trip, Matthew will travel to the remote rural villages where he was raised, in the mountainous, forested Nkongho-Mbo region, and collect at least 100 samples from three villages, which will then be sent to a lab to be screened for A00. More in-depth testing will focus on those A00 samples.
Planning and discussions are underway regarding which labs, probably more than one, will perform both SNP and STR testing on the samples. A full Y-chromosome sequence is on our wish list. This fundraising campaign, our first, will be limited to funding the DNA collection fieldwork. We'll be asking for your donations for lab testing in a separate campaign, in the near future. We need to limit the total amount of each fundraising campaign's goal to a modest amount we have some confidence of achieving within the set time, due to the all-or-nothing system used by Microryza.A00 is separated from the rest of mankind by >200 thousand years, but how closely related are different A00 chromosomes? It is necessary to have multiple samples to attempt to answer that question; I think it is quite probable that A00 is not limited to the area of Cameroon where it was discovered, but it probably makes sense to focus on it, since it is most likely to yield additional A00 samples given a sample size of 100.
A full Y-chromosome sequence of an A00 chromosome would also be very useful, since it would allow us to estimate its divergence from the A0-T remainder of mankind more securely.
The $2,500 they are seeking to launch their first field trip seems like a bargain to me.
Y chromosomes in Lingayat and Vokkaliga Dravidians from SW India

The virtual absence of Z283 subclades, namely Z280 and M458, and the total representation of R1a1a-derived samples by the Z93 marker in our dataset support an earlier observation that the M198 chromosome likely differentiated in the region between Eastern Europe and South Asia (Pamjav et al., 2012), and subsequently expanded in opposite directions. However, it will require additional R1a1a* samples from different populations across Eurasia to comprehensively evaluate the geographic origins, distribution and ethno-linguistic associations of the individual M198-derived lineages (Pamjav et al., 2012).This study extends the results of Pamjav et al. 2012 which found only Z93 within R1a1 in mainland Indian populations. The authors estimate 12.8ky as the age of R-Z93 in the Lingayat, but since this uses the evolutionary mutation rate it should actually be divided by a factor of 3.6 which translates into ~1,500BC. So, it seems quite likely that R-Z93 moved from Central->South Asia during the Bronze Age, both on account of its age and the fact that it is a subset of Central Asian diversity. Haplogroup R2 with a nominal age of ~22ky in the Lingayat seems more like a Neolithic lineage.
Also of interest:
Another haplogroup that is associated with the spread of agriculture from the Fertile Crescent and Anatolia regions is J2-M172 (Cinnioğlu et al., 2004 and Semino et al., 2004). According to Sahoo et al. (2006), only J2 lineages, originating from West Asia rather than Central Asia, represent an external contribution to the Indian paternal gene pool. In particular, subclade J2a-M410 is believed to have entered through the northwestern corridor and subsequently diffused to the south and east (Sahoo et al., 2006 and Thangaraj et al., 2010). This haplogroup is present exclusively in the Lingayat (6.93%), except for one individual from Vokkaliga, suggesting gene flow from West Asia (Sahoo et al., 2006 and Thangaraj et al., 2010). Interestingly, four J2b2-M241 Lingayat males displayed a null allele at DYS458 and failed to produce the AMGY PCR amplicon while their X homolog (AMGX) amplified successfully. Comparison of Y-STR haplotypes of the affected males from the present study with those from the literature (Cadenas et al., 2006), demonstrated a high level of allele sharing, implying shared paternal lineages or a recent common ancestry for these groups of individuals.According to the paper the Lingayat are a community that originally attracted members from across the caste system, while the Vokkaliga are involved in farming. An uneven distribution of haplogroup J2a has been previously observed, so I guess this paper adds to this evidence.
Gene Available online 7 May 2013
Indigenous and foreign Y-chromosomes characterize the Lingayat and Vokkaliga populations of Southwest India
Shilpa Chennakrishnaiah et al.
Previous studies have shown that India's vast coastal rim played an important role in the dispersal of modern humans out of Africa but the Karnataka state, which is located on the southwest coast of India, remains poorly characterized genetically. In the present study, two Dravidian populations, namely Lingayat (N = 101) and Vokkaliga (N = 102), who represent the two major communities of the Karnataka state, were examined using high-resolution analyses of Y-chromosome single nucleotide polymorphisms (Y-SNPs) and seventeen short tandem repeat (Y-STR) loci. Our results revealed that the majority of the Lingayat and Vokkaliga paternal gene pools are composed of four Y-chromosomal haplogroups (H, L, F* and R2) that are frequent in the Indian subcontinent. The high level of L1-M76 chromosomes in the Vokkaligas suggests an agricultural expansion in the region, while the predominance of R1a1a1b2-Z93 and J2a-M410 lineages in the Lingayat indicates gene flow from neighboring south Indian populations and West Asia, respectively. Lingayat (0.9981) also exhibits a relatively high haplotype diversity compared to Vokkaliga (0.9901), supporting the historical record that the Lingayat originated from multiple source populations. In addition, we detected ancient lineages such as F*-M213, H*-M69 and C*-M216 that may be indicative of genetic signatures of the earliest settlers who reached India after their migration out of Africa.
Link
July 21, 2013
Fst paper + EIGENSOFT 5.0
Razib covers a new paper on the varieties of Fst and its dependence on rare variants; this should be a very useful read for anyone interested in this widely used measure of genetic differentiation.
There is also a new 5.0 release of the EIGENSOFT suite of applications which can do (among other things) Fst. From the README file:
Genome Research doi: 10.1101/gr.154831.113
Estimating and interpreting Fst: the impact of rare variants
Gaurav Bhatia et al.
In a pair of seminal papers, Sewall Wright and Gustave Malécot introduced FST as a measure of structure in natural populations. In the decades that followed, a number of papers provided differing definitions, estimation methods, and interpretations beyond Wright's. While this diversity in methods has enabled many studies in genetics, it has also introduced confusion about how to estimate FST from available data. Considering this confusion, wide variation in published estimates of FST for pairs of HapMap populations is a cause for concern. These estimates changed- in some cases more than two-fold- when comparing estimates from genotyping arrays to those from sequence data (1000 Genomes Project Consortium 2010; International HapMap 3 Consortium 2010). Indeed, changes in FST from sequencing data might be expected data due to population genetic factors affecting rare variants. While rare variants do influence the result, we show that this is largely through differences in estimation methods. Correcting for this yields estimates of FST that are much more concordant between sequence and genotype data. These differences relate to three specific issues: (1) estimating FST for a single SNP, (2) combining estimates of FST across multiple SNPs, and (3) selecting the set of SNPs used in the computation. Changes in each of these aspects of estimation may result in FST estimates that are highly divergent from one another. Here, we clarify these issues and propose solutions.
Link
There is also a new 5.0 release of the EIGENSOFT suite of applications which can do (among other things) Fst. From the README file:
NEW features of EIGENSOFT version 5.0 include (see POPGEN/README):
-- New option lsqproject for PCA projection with large amounts of missing data
-- New options grmoutname and grmbinary to output genetic relationship matrix,
compatible with GCTA software (v1.13)
-- Expanded options for LD regression in computing genetic relationship matrix
-- Bug fix for PLINK format files with out-of-order SNPs
NOTE: multi-threading is no longer supported. Users wishing to run in multi-thre
aded
mode are recommended to use EIG4.2
NOTE: fortran compiler is no longer required to build EIGENSOFT. (However, the
lapack and blas libraries must still be installed on the system)
Genome Research doi: 10.1101/gr.154831.113
Estimating and interpreting Fst: the impact of rare variants
Gaurav Bhatia et al.
In a pair of seminal papers, Sewall Wright and Gustave Malécot introduced FST as a measure of structure in natural populations. In the decades that followed, a number of papers provided differing definitions, estimation methods, and interpretations beyond Wright's. While this diversity in methods has enabled many studies in genetics, it has also introduced confusion about how to estimate FST from available data. Considering this confusion, wide variation in published estimates of FST for pairs of HapMap populations is a cause for concern. These estimates changed- in some cases more than two-fold- when comparing estimates from genotyping arrays to those from sequence data (1000 Genomes Project Consortium 2010; International HapMap 3 Consortium 2010). Indeed, changes in FST from sequencing data might be expected data due to population genetic factors affecting rare variants. While rare variants do influence the result, we show that this is largely through differences in estimation methods. Correcting for this yields estimates of FST that are much more concordant between sequence and genotype data. These differences relate to three specific issues: (1) estimating FST for a single SNP, (2) combining estimates of FST across multiple SNPs, and (3) selecting the set of SNPs used in the computation. Changes in each of these aspects of estimation may result in FST estimates that are highly divergent from one another. Here, we clarify these issues and propose solutions.
Link
July 18, 2013
81-100 thousand year old modern humans from China
From the paper:
Such early dates are often seen as conflicting with the molecular clock (e.g., West and East Eurasians don't appear to have diverged ~100 thousand years ago no matter what assumptions about mutation rate one makes). But, we should not forget that divergence times can be suppressed either due to (i) admixture between the divergent populations, or (ii) due to an expansion of a highly successful population.
Journal of Human Evolution doi:10.1016/j.jhevol.2013.05.002
Mass spectrometric U-series dating of Huanglong Cave in Hubei Province, central China: Evidence for early presence of modern humans in eastern Asia
Guanjun Shen et al.
Most researchers believe that anatomically modern humans (AMH) first appeared in Africa 160-190 ka ago, and would not have reached eastern Asia until ∼50 ka ago. However, the credibility of these scenarios might have been compromised by a largely inaccurate and compressed chronological framework previously established for hominin fossils found in China. Recently there has been a growing body of evidence indicating the possible presence of AMH in eastern Asia ca. 100 ka ago or even earlier. Here we report high-precision mass spectrometric U-series dating of intercalated flowstone samples from Huanglong Cave, a recently discovered Late Pleistocene hominin site in northern Hubei Province, central China. Systematic excavations there have led to the in situ discovery of seven hominin teeth and dozens of stone and bone artifacts. The U-series dates on localized thin flowstone formations bracket the hominin specimens between 81 and 101 ka, currently the most narrow time span for all AMH beyond 45 ka in China, if the assignment of the hominin teeth to modern Homo sapiens holds. Alternatively this study provides further evidence for the early presence of an AMH morphology in China, through either independent evolution of local archaic populations or their assimilation with incoming AMH. Along with recent dating results for hominin samples from Homo erectus to AMH, a new extended and continuous timeline for Chinese hominin fossils is taking shape, which warrants a reconstruction of human evolution, especially the origins of modern humans in eastern Asia.
Link
The presumption of a late appearance of AMH in eastern Asia has been largely prompted by a remarkable yet dubious ‘gap’ between 100 and 40 ka ago lacking any human fossils, specifically between the latest archaic H. sapiens (i.e., Xujiayao, 104–125 ka and Maba, 129–135 ka) and the earliest modern H. sapiens (i.e., Ziyang, 35-40 ka) (data pooled from Wu and Poirier, 1995). In addition, genetic studies of present-day Chinese populations have supported the late appearance of AMH in eastern Asia (e.g., Chu et al., 1998). From this perspective, the AMH fossils from Tianyuan Cave at Zhoukoudian in northern China, dated to 39-42 ka B.P. (Before Present), have been readily accepted as the earliest representatives of modern H. sapiens in China, and the earliest evidence for the influx of African genes into the indigenous population (e.g., Shang et al., 2007; Cartmill and Smith, 2009; Endicott et al., 2009).
However, the credibility of this supposed ~60 ka gap in the hominin fossil record and a late appearance of modern humans in China might have been compromised by the exclusion of a number of hominin fossils claimed to represent AMH from various limestone caves in southern China with more ancient dates, including Liujiang (between 68 and 153 ka, and most probably between 111 and 139 ka, Shen et al., 2002b), Ganqian (94–220 ka, Shen et al., 2002a), Bailiandong (>160 ka, Shen et al., 2001b), and Zhirendong (>100 ka, Liu et al., 2010a) in Guangxi. Based on our work on the sites of H. erectus and of both archaic and modern H. sapiens over the past twenty plus years, we argue that the temporal framework in China has been artificially ‘compressed and gapped,’ meaning that due to limitations in previous dating techniques and practices, the ages of Chinese hominin fossils have been significantly postdated (compressed), and that a temporal gap between archaic H. sapiens and AMH has been artificially created (gapped). To this scenario of the possible early presence of modern humans in China, here we report evidence from Huanglong Cave, a recently discovered Late Pleistocene hominin fossil- and stone artifact-bearing site, and discuss its impact on the mode of recent human evolution in eastern Asia.I guess a lot depends on whether these teeth are accepted as belonging to AMH; if this holds, the early range of AMH must need to be extended even further east.
Such early dates are often seen as conflicting with the molecular clock (e.g., West and East Eurasians don't appear to have diverged ~100 thousand years ago no matter what assumptions about mutation rate one makes). But, we should not forget that divergence times can be suppressed either due to (i) admixture between the divergent populations, or (ii) due to an expansion of a highly successful population.
Journal of Human Evolution doi:10.1016/j.jhevol.2013.05.002
Mass spectrometric U-series dating of Huanglong Cave in Hubei Province, central China: Evidence for early presence of modern humans in eastern Asia
Guanjun Shen et al.
Most researchers believe that anatomically modern humans (AMH) first appeared in Africa 160-190 ka ago, and would not have reached eastern Asia until ∼50 ka ago. However, the credibility of these scenarios might have been compromised by a largely inaccurate and compressed chronological framework previously established for hominin fossils found in China. Recently there has been a growing body of evidence indicating the possible presence of AMH in eastern Asia ca. 100 ka ago or even earlier. Here we report high-precision mass spectrometric U-series dating of intercalated flowstone samples from Huanglong Cave, a recently discovered Late Pleistocene hominin site in northern Hubei Province, central China. Systematic excavations there have led to the in situ discovery of seven hominin teeth and dozens of stone and bone artifacts. The U-series dates on localized thin flowstone formations bracket the hominin specimens between 81 and 101 ka, currently the most narrow time span for all AMH beyond 45 ka in China, if the assignment of the hominin teeth to modern Homo sapiens holds. Alternatively this study provides further evidence for the early presence of an AMH morphology in China, through either independent evolution of local archaic populations or their assimilation with incoming AMH. Along with recent dating results for hominin samples from Homo erectus to AMH, a new extended and continuous timeline for Chinese hominin fossils is taking shape, which warrants a reconstruction of human evolution, especially the origins of modern humans in eastern Asia.
Link
July 17, 2013
Sex assignment of ancient samples from shotgun sequencing data

Interestingly, many of the ancient European samples that appeared over the last couple of years turn out to be male, and so do some Neandertals; the latter might be especially important, because the available high-coverage Neandertal genome appears to be a female; while we do know about the divergence of modern human and Neandertal autosomal DNA and mtDNA, we have no clue, at the moment about the Y-chromosome MRCA of modern humans and Neandertals; it's not clear to me how good the DNA from the Neandertal males.
A version of the paper has been posted here.
Journal of Archaeological Science Available online 14 July 2013
Accurate sex identification of ancient human remains using DNA shotgun sequencing
Pontus Skoglund et al.
Accurate identification of the biological sex of ancient remains is vital for critically testing hypotheses about social structure in prehistoric societies. However, morphological methods are imprecise for juvenile individuals and fragmentary remains, and molecular methods that rely on particular sex-specific marker loci such as the amelogenin gene suffer from allelic dropout and sensitivity to modern contamination. Analyzing shotgun sequencing data from 14 present-day humans of known biological sex and 16 ancient individuals from a time span of 100 to ~70,000 years ago, we show that even relatively sparse shotgun sequencing (about 100,000 human sequences) can be used to reliably identify chromosomal sex simply by considering the ratio of sequences aligning to the X and Y chromosomes, and highlight two examples where the genetic assignments indicate morphological misassignment. Furthermore, we show that accurate sex identification of highly degraded remains can be performed in the presence of substantial amounts of present-day contamination by utilizing the signature of cytosine deamination, a characteristic feature of ancient DNA.
Link
Inbreeding and cognitive ability
From the paper:
Genome-wide estimates of inbreeding in unrelated individuals and their association with cognitive ability
Robert A Power et al.
The consequence of reduced cognitive ability from inbreeding has long been investigated, mainly restricted to cousin–cousin marriages. Molecular genetic techniques now allow us to test the relationship between increased ancestral inbreeding and cognitive ability in a population of traditionally unrelated individuals. In a representative UK sample of 2329 individuals, we used genome-wide SNP data to estimate the percentage of the genome covered by runs of homozygous SNPs (ROH). This was tested for association with general cognitive ability, as well as measures of verbal and non-verbal ability. Further, association was tested between these traits and specific ROH. Burden of ROH was not associated with cognitive ability after correction for multiple testing, although burden of ROH was nominally associated with increased non-verbal cognitive ability (P=0.03). Moreover, although no individual ROH was significantly associated with cognitive ability, there was a significant bias towards increased cognitive ability in carriers of ROH (P=0.002). A potential explanation for these results is increased positive assortative mating in spouses with higher cognitive ability, although we found no evidence in support of this hypothesis in a separate sample. Reduced minor allele frequency across the genome was associated with higher cognitive ability, which could contribute to an apparent increase in ROH. This may reflect minor alleles being more likely to be deleterious.
Link
Our results show that within a representative UK population sample there was a weak nominally significant association between burden of autosomal runs of homozygosity and higher non-verbal cognitive ability. This nominal association with increased cognitive ability is counterintuitive when compared with the results from more extreme inbreeding based on pedigree information.1, 2, 3 A potential explanation for this direction of effect is that individuals with higher cognitive ability might show greater positive assortative mating, which would lead to increased homozygosity at loci for higher cognitive ability in their offspring. However, in a separate sample we showed that greater positive assortative mating was not associated with higher cognitive ability. While these findings seem to provide clear evidence against this hypothesis, it is possible that the genome-wide genetic findings reflect historical mating habits that no longer exist today. It should also be noted that there was a reduction in the standard deviations for spousal correlations in the increased cognitive ability groups by an average of 6% compared with the decreased cognitive ability group (see Table 3), which could reflect lesser genetic variability in the high ability couples or a ceiling effect on the cognitive tests. This lesser phenotypic variability at the high ability end would have a small effect in reducing the spouse correlations and potentially confound our analysis.
Genetic variants were found to have a slight (though significant) reduction in minor allele frequency across the genome in individuals in the top quartile of general cognitive ability compared with those in the bottom quartile (means of 21823 and 21824), which in turn could lead to increased homozygosity by chance. This could indicate that these individuals descend from subtly different ancestral populations that experienced loss of variation. This difference in ancestry may be correlated with either genetic variants for improved cognitive ability, or with social or environmental backgrounds that lead to higher cognitive ability, though our analysis of FROH corrected for socio-economic status and population stratification. Another potential explanation is that the reduced minor allele frequency in the high cognitive ability is reflective of the less frequent allele being deleterious due to selection against it. As a result, these high-functioning individuals could benefit from having more major alleles at fixation and a reduced burden of rarer deleterious variants.
Overall, these results highlight the importance of understanding mating habits, such as inbreeding and assortative mating, when investigating the genetic architecture of complex traits such as cognitive ability. The results certainly suggest that there is no large effect of FROH on reduced cognitive ability, the expected direction of effect. The nominally significant associations found in this study may even suggest that in the case of non-verbal cognitive ability, beneficial associations with homozygosity at specific loci might outweigh the negative effects of genome-wide inbreeding and that the relationship between inbreeding and cognitive ability may be more complicated than previously thought.European Journal of Human Genetics , (17 July 2013) | doi:10.1038/ejhg.2013.155
Genome-wide estimates of inbreeding in unrelated individuals and their association with cognitive ability
Robert A Power et al.
The consequence of reduced cognitive ability from inbreeding has long been investigated, mainly restricted to cousin–cousin marriages. Molecular genetic techniques now allow us to test the relationship between increased ancestral inbreeding and cognitive ability in a population of traditionally unrelated individuals. In a representative UK sample of 2329 individuals, we used genome-wide SNP data to estimate the percentage of the genome covered by runs of homozygous SNPs (ROH). This was tested for association with general cognitive ability, as well as measures of verbal and non-verbal ability. Further, association was tested between these traits and specific ROH. Burden of ROH was not associated with cognitive ability after correction for multiple testing, although burden of ROH was nominally associated with increased non-verbal cognitive ability (P=0.03). Moreover, although no individual ROH was significantly associated with cognitive ability, there was a significant bias towards increased cognitive ability in carriers of ROH (P=0.002). A potential explanation for these results is increased positive assortative mating in spouses with higher cognitive ability, although we found no evidence in support of this hypothesis in a separate sample. Reduced minor allele frequency across the genome was associated with higher cognitive ability, which could contribute to an apparent increase in ROH. This may reflect minor alleles being more likely to be deleterious.
Link
July 15, 2013
Early European farmers used manure
PNAS doi: 10.1073/pnas.1305918110
Crop manuring and intensive land management by Europe’s first farmers
Amy Bogaard et al.
The spread of farming from western Asia to Europe had profound long-term social and ecological impacts, but identification of the specific nature of Neolithic land management practices and the dietary contribution of early crops has been problematic. Here, we present previously undescribed stable isotope determinations of charred cereals and pulses from 13 Neolithic sites across Europe (dating ca. 5900–2400 cal B.C.), which show that early farmers used livestock manure and water management to enhance crop yields. Intensive manuring inextricably linked plant cultivation and animal herding and contributed to the remarkable resilience of these combined practices across diverse climatic zones. Critically, our findings suggest that commonly applied paleodietary interpretations of human and herbivore δ15N values have systematically underestimated the contribution of crop-derived protein to early farmer diets.
Link
Crop manuring and intensive land management by Europe’s first farmers
Amy Bogaard et al.
The spread of farming from western Asia to Europe had profound long-term social and ecological impacts, but identification of the specific nature of Neolithic land management practices and the dietary contribution of early crops has been problematic. Here, we present previously undescribed stable isotope determinations of charred cereals and pulses from 13 Neolithic sites across Europe (dating ca. 5900–2400 cal B.C.), which show that early farmers used livestock manure and water management to enhance crop yields. Intensive manuring inextricably linked plant cultivation and animal herding and contributed to the remarkable resilience of these combined practices across diverse climatic zones. Critically, our findings suggest that commonly applied paleodietary interpretations of human and herbivore δ15N values have systematically underestimated the contribution of crop-derived protein to early farmer diets.
Link
July 12, 2013
A Middle Paleolithic link between North Africa and the Thar desert

And, one more piece of evidence against the idea that early modern humans in Eurasia died out and were replaced wholesale circa 50-60 thousand years ago from a fresh OoA impulse. It would have been possible to suppose such a thing if the evidence for MP African influence was minor or geographically localized, but much more difficult when it extends over a wide region.
Of course, the attribution of the Katoati assemblages to modern humans is done indirectly by linking them to MSA sites of the Sahara, but the proliferation of real archaeological sites (see map) that can be linked to OoA makes it difficult to adopt the idea of an archaeologically invisible late OoA that (somehow) replaced all previous inhabitants.
Quaternary Science Reviews Available online 5 July 2013
Middle Palaeolithic occupation in the Thar Desert during the Upper Pleistocene: the signature of a modern human exit out of Africa?
James Blinkhorn et al.
The Thar Desert marks the transition from the Saharo-Arabian deserts to the Oriental biogeographical zone and is therefore an important location in understanding hominin occupation and dispersal during the Upper Pleistocene. Here, we report the discovery of stratified Middle Palaeolithic assemblages at Katoati in the north-eastern Thar Desert, dating to Marine Isotope Stages (MIS) 5 and the MIS 4–3 boundary, during periods of enhanced humidity. Hominins procured cobbles from gravels at the site as evidenced by early stages of stone tool reduction, with a component of more formalised point production. The MIS 5c assemblages at Katoati represent the earliest securely dated Middle Palaeolithic occupation of South Asia. Distinctive artefacts identified in both MIS 5 and MIS 4–3 boundary horizons match technological entities observed in Middle Palaeolithic assemblages in South Asia, Arabia and Middle Stone Age sites in the Sahara. The evidence from Katoati is consistent with arguments for the dispersal of Homo sapiens populations from Africa across southern Asia using Middle Palaeolithic technologies.
Link
July 11, 2013
Native Native American dogs
Proc. R. Soc. B doi: 10.1098/rspb.2013.1142
Pre-Columbian origins of Native American dog breeds, with only limited replacement by European dogs, confirmed by mtDNA analysis
Barbara van Asch et al.
Dogs were present in pre-Columbian America, presumably brought by early human migrants from Asia. Studies of free-ranging village/street dogs have indicated almost total replacement of these original dogs by European dogs, but the extent to which Arctic, North and South American breeds are descendants of the original population remains to be assessed. Using a comprehensive phylogeographic analysis, we traced the origin of the mitochondrial DNA lineages for Inuit, Eskimo and Greenland dogs, Alaskan Malamute, Chihuahua, xoloitzcuintli and perro sín pelo del Peru, by comparing to extensive samples of East Asian (n = 984) and European dogs (n = 639), and previously published pre-Columbian sequences. Evidence for a pre-Columbian origin was found for all these breeds, except Alaskan Malamute for which results were ambigous. No European influence was indicated for the Arctic breeds Inuit, Eskimo and Greenland dog, and North/South American breeds had at most 30% European female lineages, suggesting marginal replacement by European dogs. Genetic continuity through time was shown by the sharing of a unique haplotype between the Mexican breed Chihuahua and ancient Mexican samples. We also analysed free-ranging dogs, confirming limited pre-Columbian ancestry overall, but also identifying pockets of remaining populations with high proportion of indigenous ancestry, and we provide the first DNA-based evidence that the Carolina dog, a free-ranging population in the USA, may have an ancient Asian origin.
Link
Pre-Columbian origins of Native American dog breeds, with only limited replacement by European dogs, confirmed by mtDNA analysis
Barbara van Asch et al.
Dogs were present in pre-Columbian America, presumably brought by early human migrants from Asia. Studies of free-ranging village/street dogs have indicated almost total replacement of these original dogs by European dogs, but the extent to which Arctic, North and South American breeds are descendants of the original population remains to be assessed. Using a comprehensive phylogeographic analysis, we traced the origin of the mitochondrial DNA lineages for Inuit, Eskimo and Greenland dogs, Alaskan Malamute, Chihuahua, xoloitzcuintli and perro sín pelo del Peru, by comparing to extensive samples of East Asian (n = 984) and European dogs (n = 639), and previously published pre-Columbian sequences. Evidence for a pre-Columbian origin was found for all these breeds, except Alaskan Malamute for which results were ambigous. No European influence was indicated for the Arctic breeds Inuit, Eskimo and Greenland dog, and North/South American breeds had at most 30% European female lineages, suggesting marginal replacement by European dogs. Genetic continuity through time was shown by the sharing of a unique haplotype between the Mexican breed Chihuahua and ancient Mexican samples. We also analysed free-ranging dogs, confirming limited pre-Columbian ancestry overall, but also identifying pockets of remaining populations with high proportion of indigenous ancestry, and we provide the first DNA-based evidence that the Carolina dog, a free-ranging population in the USA, may have an ancient Asian origin.
Link
July 10, 2013
Population history of middle Euphrates valley
HOMO - Journal of Comparative Human Biology
Available online 3 July 2013
Population history of the middle Euphrates valley: Dental non-metric traits at Tell Ashara, Tell Masaikh and Jebel Mashtale, Syria
Arkadiusz Sołtysiak, Marta Bialon
Fifty-nine dental non-metric traits were scored using Arizona State University Dental Anthropology System on a sample of teeth from 350 human skeletons excavated at three sites in the lower middle Euphrates valley. The dataset was divided into six chronological subsets: Early Bronze Age, Middle Bronze Age, Early Iron Age with Neo-Assyrian period, Classical/Late Antiquity, Early Islamic (Umayyad and Abbasid) period and Modern period. The matrix of Mean Measure of Divergence values exhibited temporal homogeneity of the sample with only dental non-metric trait scores in the Modern subset differing significantly from most other subsets. Such a result suggests that no major gene flow occurred in the middle Euphrates valley between the 3rd millennium BCE and the early 2nd millennium CE. Only after the Mongolian invasion and large depopulation of northern Mesopotamia in the 13th century CE a major population change occurred when the area was taken over in the 17th century by Bedouin tribes from the Arabian Peninsula.
Link
Population history of the middle Euphrates valley: Dental non-metric traits at Tell Ashara, Tell Masaikh and Jebel Mashtale, Syria
Arkadiusz Sołtysiak, Marta Bialon
Fifty-nine dental non-metric traits were scored using Arizona State University Dental Anthropology System on a sample of teeth from 350 human skeletons excavated at three sites in the lower middle Euphrates valley. The dataset was divided into six chronological subsets: Early Bronze Age, Middle Bronze Age, Early Iron Age with Neo-Assyrian period, Classical/Late Antiquity, Early Islamic (Umayyad and Abbasid) period and Modern period. The matrix of Mean Measure of Divergence values exhibited temporal homogeneity of the sample with only dental non-metric trait scores in the Modern subset differing significantly from most other subsets. Such a result suggests that no major gene flow occurred in the middle Euphrates valley between the 3rd millennium BCE and the early 2nd millennium CE. Only after the Mongolian invasion and large depopulation of northern Mesopotamia in the 13th century CE a major population change occurred when the area was taken over in the 17th century by Bedouin tribes from the Arabian Peninsula.
Link
July 08, 2013
Continuity of microblade technology in India since 45ka
An interesting new paper extends continuity of microblade technologies in India to ~45ka, and hence makes it probable that these were introduced by AMH together with the UP colonization of the rest of Eurasia.
I am not sure that the authors' suggestion that early modern humans were "tropically adapted" is certain. Personally, my idea du jour is to derive them from the Saharo-Arabian belt. In any case, as an advocate of "early OOA" (in the sense of pre-UP/LSA), it makes sense to me that modern humans in Eurasia would be initially climate-limited and at a disadvantage vis a vis archaic Eurasians inhabiting regions for which they were maladapted. In my opinion, it was the technological revolution of ~50ka being responsible for the extension of their range at the expense of other Eurasians.
PLoS ONE 8(7): e69280. doi:10.1371/journal.pone.0069280
Continuity of Microblade Technology in the Indian Subcontinent Since 45 ka: Implications for the Dispersal of Modern Humans
Sheila Mishra et al.
We extend the continuity of microblade technology in the Indian Subcontinent to 45 ka, on the basis of optical dating of microblade assemblages from the site of Mehtakheri, (22° 13' 44″ N Lat 76° 01' 36″ E Long) in Madhya Pradesh, India. Microblade technology in the Indian Subcontinent is continuously present from its first appearance until the Iron Age (~3 ka), making its association with modern humans undisputed. It has been suggested that microblade technology in the Indian Subcontinent was developed locally by modern humans after 35 ka. The dates reported here from Mehtakheri show this inference to be untenable and suggest alternatively that this technology arrived in the Indian Subcontinent with the earliest modern humans. It also shows that modern humans in Indian Subcontinent and SE Asia were associated with differing technologies and this calls into question the “southern dispersal” route of modern humans from Africa through India to SE Asia and then to Australia. We suggest that modern humans dispersed from Africa in two stages coinciding with the warmer interglacial conditions of MIS 5 and MIS 3. Competitive interactions between African modern humans and Indian archaics who shared an adaptation to tropical environments differed from that between modern humans and archaics like Neanderthals and Denisovans, who were adapted to temperate environments. Thus, while modern humans expanded into temperate regions during warmer climates, their expansion into tropical regions, like the Indian Subcontinent, in competition with similarly adapted populations, occurred during arid climates. Thus modern humans probably entered the Indian Subcontinent during the arid climate of MIS 4 coinciding with their disappearance from the Middle East and Northern Africa. The out of phase expansion of modern humans into tropical versus temperate regions has been one of the factors affecting the dispersal of modern humans from Africa during the period 200–40 ka.
Link
I am not sure that the authors' suggestion that early modern humans were "tropically adapted" is certain. Personally, my idea du jour is to derive them from the Saharo-Arabian belt. In any case, as an advocate of "early OOA" (in the sense of pre-UP/LSA), it makes sense to me that modern humans in Eurasia would be initially climate-limited and at a disadvantage vis a vis archaic Eurasians inhabiting regions for which they were maladapted. In my opinion, it was the technological revolution of ~50ka being responsible for the extension of their range at the expense of other Eurasians.
PLoS ONE 8(7): e69280. doi:10.1371/journal.pone.0069280
Continuity of Microblade Technology in the Indian Subcontinent Since 45 ka: Implications for the Dispersal of Modern Humans
Sheila Mishra et al.
We extend the continuity of microblade technology in the Indian Subcontinent to 45 ka, on the basis of optical dating of microblade assemblages from the site of Mehtakheri, (22° 13' 44″ N Lat 76° 01' 36″ E Long) in Madhya Pradesh, India. Microblade technology in the Indian Subcontinent is continuously present from its first appearance until the Iron Age (~3 ka), making its association with modern humans undisputed. It has been suggested that microblade technology in the Indian Subcontinent was developed locally by modern humans after 35 ka. The dates reported here from Mehtakheri show this inference to be untenable and suggest alternatively that this technology arrived in the Indian Subcontinent with the earliest modern humans. It also shows that modern humans in Indian Subcontinent and SE Asia were associated with differing technologies and this calls into question the “southern dispersal” route of modern humans from Africa through India to SE Asia and then to Australia. We suggest that modern humans dispersed from Africa in two stages coinciding with the warmer interglacial conditions of MIS 5 and MIS 3. Competitive interactions between African modern humans and Indian archaics who shared an adaptation to tropical environments differed from that between modern humans and archaics like Neanderthals and Denisovans, who were adapted to temperate environments. Thus, while modern humans expanded into temperate regions during warmer climates, their expansion into tropical regions, like the Indian Subcontinent, in competition with similarly adapted populations, occurred during arid climates. Thus modern humans probably entered the Indian Subcontinent during the arid climate of MIS 4 coinciding with their disappearance from the Middle East and Northern Africa. The out of phase expansion of modern humans into tropical versus temperate regions has been one of the factors affecting the dispersal of modern humans from Africa during the period 200–40 ka.
Link
July 05, 2013
Early agriculture from Iran
A quite useful map from the press release of this paper:

The following map from an accompanying perspective is also quite interesting; "Dates in blue denote early cultivation of wild cereals" but "Ongoing excavations in central Anatolia and Cyprus are pushing dates back in these areas."

The issue of whether there was a single or (more likely multiple) areas of early agriculture is potentially important as it would imply that there were genetically differentiated (due to geographic distance) populations in the Neolithic womb of nations. In a global, or even a Eurasian context, these populations would be relatively genetically close, but not identical; it would be interesting to see to what extent present-day differentiation in the Near East reflects those early differences as opposed to more recent events.
Science 5 July 2013: Vol. 341 no. 6141 pp. 65-67
Emergence of Agriculture in the Foothills of the Zagros Mountains of Iran
Simone Riehl et al.
The role of Iran as a center of origin for domesticated cereals has long been debated. High stratigraphic resolution and rich archaeological remains at the aceramic Neolithic site of Chogha Golan (Ilam Province, present-day Iran) reveal a sequence ranging over 2200 years of cultivation of wild plants and the first appearance of domesticated-type species. The botanical record from Chogha Golan documents how the inhabitants of the site cultivated wild barley (Hordeum spontaneum) and other wild progenitor species of modern crops, such as wild lentil and pea. Wild wheat species (Triticum spp.) are initially present at less than 10% of total plant species but increase to more than 20% during the last 300 years of the sequence. Around 9800 calendar years before the present, domesticated-type emmer appears. The archaeobotanical remains from Chogha Golan represent the earliest record of long-term plant management in Iran.
Link

The following map from an accompanying perspective is also quite interesting; "Dates in blue denote early cultivation of wild cereals" but "Ongoing excavations in central Anatolia and Cyprus are pushing dates back in these areas."
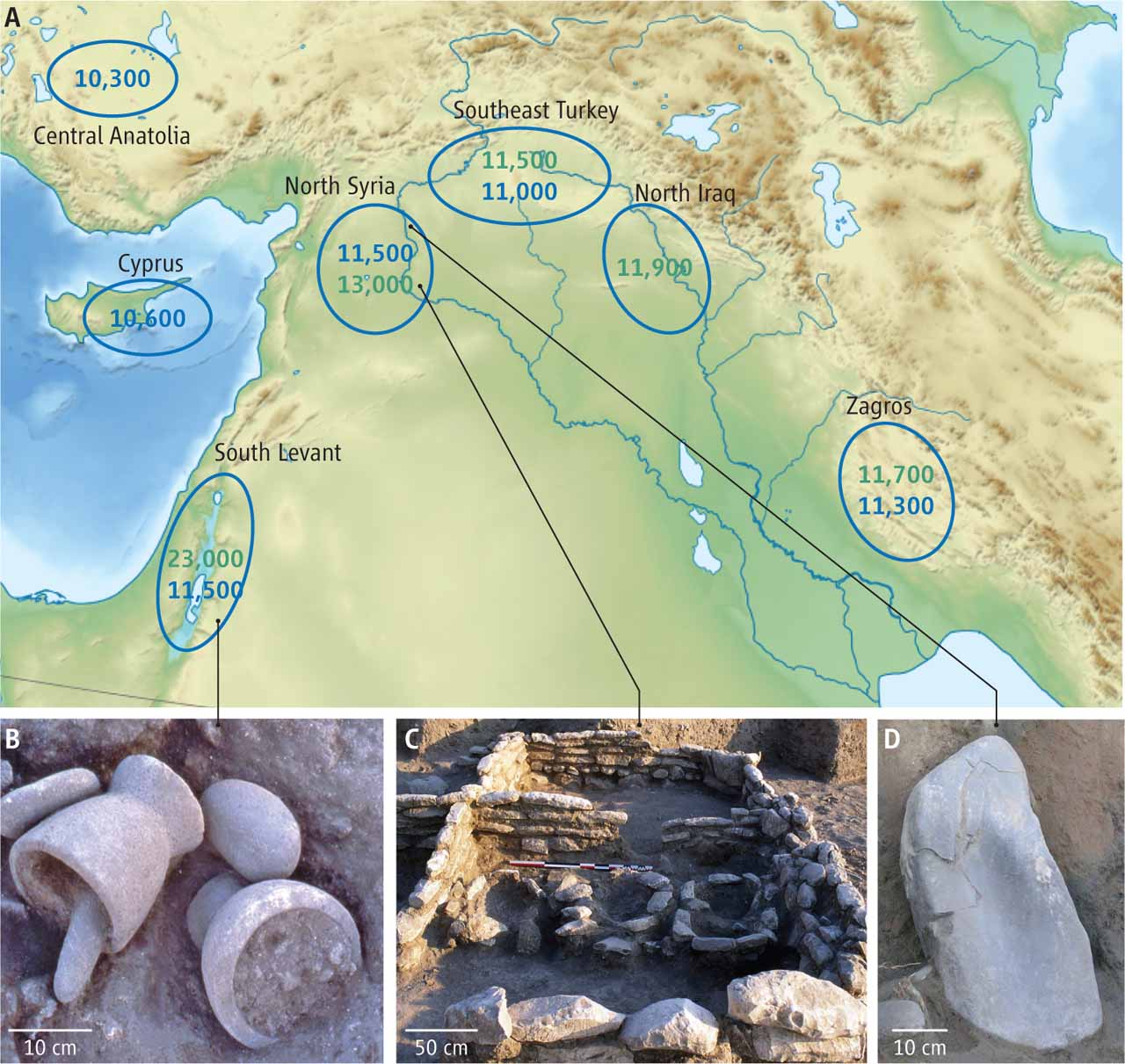
The issue of whether there was a single or (more likely multiple) areas of early agriculture is potentially important as it would imply that there were genetically differentiated (due to geographic distance) populations in the Neolithic womb of nations. In a global, or even a Eurasian context, these populations would be relatively genetically close, but not identical; it would be interesting to see to what extent present-day differentiation in the Near East reflects those early differences as opposed to more recent events.
Science 5 July 2013: Vol. 341 no. 6141 pp. 65-67
Emergence of Agriculture in the Foothills of the Zagros Mountains of Iran
Simone Riehl et al.
The role of Iran as a center of origin for domesticated cereals has long been debated. High stratigraphic resolution and rich archaeological remains at the aceramic Neolithic site of Chogha Golan (Ilam Province, present-day Iran) reveal a sequence ranging over 2200 years of cultivation of wild plants and the first appearance of domesticated-type species. The botanical record from Chogha Golan documents how the inhabitants of the site cultivated wild barley (Hordeum spontaneum) and other wild progenitor species of modern crops, such as wild lentil and pea. Wild wheat species (Triticum spp.) are initially present at less than 10% of total plant species but increase to more than 20% during the last 300 years of the sequence. Around 9800 calendar years before the present, domesticated-type emmer appears. The archaeobotanical remains from Chogha Golan represent the earliest record of long-term plant management in Iran.
Link
SMBE 2013 abstracts
Some abstracts from the SMBE 2013 conference that will take place next week.
Legacy of Early Migrants in Neolithic East Asian Hunter-Gatherer from Fukushima, Japan
K.K. Hideaki et al.
Clarifying the genetic relationship between Neolithic East Asian Hunter-Gatherer, Jomon people, and modern human populations is one of the Keystones to understand the controversial history of modern East Asian populations. Jomon people inhabited in the Japanese archipelago from 16,000 years ago, and their origin and the relationship with modern humans have been debated for a long time. To solve these questions, we obtained 20 million base pairs of genomic DNA from a ~4,000-year-old Jomon male tooth, excavated in Sanganji shell mound, Fukushima, Japan. We compared his genetic components with the data of modern worldwide populations. Our major findings are: (1) Sanganji Jomon was very similar with modern East Asians when we compared the worldwide populations in the PCA plot; (2) when only East Asians were compared, Sanganji Jomon was distant from both modern Northeast and Southeast Asians, indicating that Sanganji Jomon people were already isolated from other continental populations for a long time; (3) the Sanganji Jomon male shared more SNP alleles with southern and northern minorities in China than geographically close Han Chinese, implying a complex history of people in China after the divergence between Jomon ancestors and Eurasian continent people; (4) Sanganji Jomon is genetically closer to modern mainland Japanese than continental populations, indicating that some of their components were transmitted to modern Japanese; (5) within Japanese archipelago, Ainu and Ryukyuan (the populations of northern and southern edges of the Japanese archipelago) have more Jomon components than mainland Japanese, indicating that the genetic effect of agricultural people who migrated from the Eurasian continent in and after the Yayoi period is stronger in Mainland Japanese than Ainu and Ryukyuan.
Extensive Gene Gain in Human Brain Evolution
Yong E. Zhang et al.
The genetic changes contributing to the evolution of the human brain have always attracted wide interest. A emerging consensus view is that while there have been no major patterns of genome-wide changes to the coding regions of brain-related genes, cis-regulatory changes of these genes have played a key role. Here, motivated by anecdotal studies of primate-specific genes implicated in brain function, we identified thousands of lineage-specific (primate-specific or rodent-specific) genes by mining syntenic vertebrate genomic alignments and examined the expression profile of these genes in both fetal and adult brains of human and mouse across different transcriptome profiling platforms. We found that an excess of lineage-specific genes are expressed in the early (fetal or infant) developing human brain compared with those in mouse brain. Expression data covering numerous subregions of the developing brain further demonstrate that these young genes are mainly transcribed in the neocortex. They originated in the evolutionary period during which the neocortex was expanding, suggesting the functional association of new genes with this newly evolving brain structure. Our data reveal that evolutionary change in the development of the human brain happened at the protein level by gene origination and also via evolution of regulatory networks, as hinted by the enrichment of primate-specific transcriptional regulators in our dataset. More than that, these ?ndings suggest that genomes are continually evolving in both sequence and content, eroding the conservation endowed by common ancestry. Despite increasing recognition of the importance of new genes, these genes are still seriously under-characterized in functional studies and that new gene annotation is inconsistent in current practice. We propose an integrative approach based on functional and evolutionary genomic methods to better annotate these non-conserved genes.
Recent Human Demography Impacts the Architecture of Genetic Disease in Populations but Not Individual Genetic Load
Yuval Simons et al.
Human populations have undergone dramatic changes in population sizes in the past 100,000 years, including a severe bottleneck of non-African populations and recent explosive population growth. There is currently great interest in how these demographic events may have affected the burden of deleterious mutations in individuals and the allele frequency spectrum of disease mutations in populations. Here we use population genetic models to show that--contrary to previous conjectures--recent human demography likely had very little impact on the average burden of deleterious mutations carried by individuals. This prediction is supported by exome sequence data showing that African American and European American individuals carry very similar burdens of damaging mutations. We next considered whether recent population growth has increased the importance of very rare mutations in disease. Our analysis predicts that, even given recent growth, it is unlikely that very rare mutations contribute a large fraction of disease heritability except for diseases that are largely due to strongly deleterious mutations. In summary, demographic history has dramatically impacted patterns of variation in different human populations, but these changes likely had little impact on either genetic load or on the importance of rare variants in most complex traits.
Functional and Population Genetic Analyses of a High-Coverage Neandertal Genome
Fernando Racimo et al.
We have sequenced the genome of a Neandertal from the Altai mountains in Siberia at 50-fold coverage. This Neandertal was located in the same cave as the Denisovan individual, but is phylogenetically closer to Neandertals from Western Eurasia. To avoid confusion, we call this individual the Altai Neandertal. Here we show an assessment of genome data quality and functional and population genetic comparisons with the Denisovan genome and a set of 25 high-coverage modern human genomes. We present an analysis of genes with recent changes in either the modern human lineage or the archaic human lineage (Neandertal+Denisova). We find enrichment for nonsynonymous changes in genes associated with melanosomes in the modern human lineage, and genes associated with particular muscoskeletal morphologies in the archaic human lineage. We utilize a compound deleteriousness scoring that allows us to combine a variety of conservation, regulatory and expression data to rank all modern and archaic-specific single-nucleotide changes and InDels across the genome, and predict which are those that could have been most disruptive in our evolutionary history. Furthermore, we overlap the modern-specific catalog with the top regions of a screen for selective sweeps exclusive to the modern human lineage, and observe enrichments for changes in genes related to ion channel activity, muscle contraction and membrane transport. Finally, we note an excess of ancestral alleles in the Denisovan individual relative to the Altai Neandertal individual, which is strongest at sites where modern humans are fixed derived. We develop an approximate Bayesian computation approach that allows us to test different models, and conclude that gene flow between the Denisovan individual and a more anciently diverged human lineage is most consistent with the patterns observed.
Multiple Episodes of Population Mixture in Southern African History
Joseph K. Pickrell et al.
The history of southern Africa involved interactions between indigenous hunter-gatherers and a range of populations that moved temporarily or permanently into the region. The influence of these interactions on the genetic structure of current populations remains unclear. Here, using patterns of linkage disequilibrium, we show that there are at least two admixture events in the genetic history of southern African hunter-gatherers and pastoralists: one involving populations related to Niger-Congo-speaking African populations, and one which introduced ancestry most closely related to west Eurasian (European or Middle Eastern) populations. We estimate that at least a few percent of ancestry in the Khoisan is derived from this latter admixture event, which occurred on average 1,200-1,800 years ago. We show that a similar signal of west Eurasian ancestry is present throughout eastern Africa; in particular, we also find evidence for two admixture events in the genetic history of several Kenyan, Tanzanian, and Somali populations, the earliest of which involved populations related to southern Europeans and which we date to approximately 2700 - 3300 years ago. We thus suggest that west Eurasian ancestry entered southern Africa indirectly through eastern Africa. These results demonstrate how large-scale genomic datasets can inform complex models of population movements, and highlight the genomic impact of largely uncharacterized back-to-Africa migrations in human history.
Mechanistic Models of Admixture and Approximate-Approximate-Bayesian Computation
Noah Rosenberg et al.
Investigations of the history of migrations that underlie admixed populations often use nonmechanistic admixture models rather than a modeling perspective that incorporates a population-genetic history of admixture built from first principles. We build a general model of admixture that mechanistically accounts for complex admixture processes, considering two source populations that contribute to the ancestry of a hybrid population, potentially with variable contributions across generations. For a random individual in the hybrid population at a given point in time, we study the fraction of admixture originating from a specific one of the source populations. Quite different admixture processes can produce identical mean admixture across individuals, but such processes typically produce different values for the variance of admixture. Interestingly, even without considering sex chromosomes, the variance of admixture for autosomes captures information about sex-specific migration in the contributions of the source populations to the admixed group. To perform inference under the model, we use approximate-approximate-Bayesian computation (AABC), a modification of approximateBayesian computation well-suited to estimation under complex mechanistic models that are computationally intensive to simulate. The model and inference method can contribute to an understanding of the theory and analysis of the history of admixed populations.Inferring Human Population History and Migration Patterns from Multiple Genome Sequences
Stephan Schiffels, Richard Durbin
The availability of human genomes from populations across the world has given rise to new inference methods that exploit high-coverage sequence data. Among the most influential recent developments is the Pairwise Sequentially Markovian Coalescenct (PSMC) by Li and Durbin, 2011. While PSMC infers the demographic history for times between 20kya and 2mya with high resolution, neither the more recent evolutionary history nor migration patterns across populations can be adressed. Here we present a new method that overcomes both of these shortcomings. The Multiple Sequentially Markovian Coalescent (MSMC) infers the recent evolutionary history within and across populations only a few thousand years ago and younger. MSMC models the pattern of mutations in multiple genome sequences under the coalescent with recombination. It fits local genealogical trees to the observed pattern, focussing on the first coalescence among any two individuals. We apply our method to the genome sequences from several family trios from the 1000 Genomes project with African, European, Asian and SouthAmerican ancestry. We infer population sizes and migration rates as a function of time with high resolution. In particular, our method resolves the more recent evolutionary history of non-African populations after the out-of-Africa event, such as the peopling of the Americas.
July 03, 2013
The history of great apes

Great ape genetic diversity and population history
Javier Prado-Martinez et al.
Most great ape genetic variation remains uncharacterized1, 2; however, its study is critical for understanding population history3, 4, 5, 6, recombination7, selection8 and susceptibility to disease9, 10. Here we sequence to high coverage a total of 79 wild- and captive-born individuals representing all six great ape species and seven subspecies and report 88.8 million single nucleotide polymorphisms. Our analysis provides support for genetically distinct populations within each species, signals of gene flow, and the split of common chimpanzees into two distinct groups: Nigeria–Cameroon/western and central/eastern populations. We find extensive inbreeding in almost all wild populations, with eastern gorillas being the most extreme. Inferred effective population sizes have varied radically over time in different lineages and this appears to have a profound effect on the genetic diversity at, or close to, genes in almost all species. We discover and assign 1,982 loss-of-function variants throughout the human and great ape lineages, determining that the rate of gene loss has not been different in the human branch compared to other internal branches in the great ape phylogeny. This comprehensive catalogue of great ape genome diversity provides a framework for understanding evolution and a resource for more effective management of wild and captive great ape populations.
Link
Origin of Early Transcaucasian Culture (aka Kura-Araxes culture)

Akhundov (2007) recently uncovered pre-Kura-Araxes/Late Chalcolithic materials from the settlement of Boyuk Kesik and the kurgan necropolis of Soyuq Bulaq in northwestern Azerbaijan, and Makharadze (2007) has also excavated a pre-Kura-Araxes kurgan, Kavtiskhevi, in central Georgia. Materials recovered from both these recent excavations can be related to remains from the metal-working Late Chalcolithic site of Leilatepe on the Karabakh steppe near Agdam (Narimanov et al. 2007) and from the earliest level at the multi-period site of Berikldeebi in Kvemo Kartli (Glonti and Dzavakhishvili 1987). They reveal the presence of early 4th millennium raised burial mounds or kurgans in the southern Caucasus. Similarly, on the basis of her survey work in eastern Anatolia north of the Oriental Taurus mountains, C. Marro (2007)likens chafffaced wares collected at Hanago in the Sürmeli Plain and Astepe and Colpan in the eastern Lake Van district in northeastern Turkey with those found at the sites mentioned above and relates these to similar wares (Amuq E/F) found south of the Taurus Mountains in northern Mesopotamia
...
The new high dating of the Maikop culture essentially signifies that there is no
chronological hiatus separating the collapse of the Chalcolithic Balkan centre of
metallurgical production and the appearance of Maikop and the sudden explosion of Caucasian metallurgical production and use of arsenical copper/bronzes. More than forty calibrated radiocarbon dates on Maikop and related materials now support this high chronology; and the revised dating for the Maikop culture means that the earliest kurgans occur in the northwestern and southern Caucasus and precede by several centuries those of the Pit-Grave (Yamnaya) cultures of the western Eurasian steppes (cf. Chernykh and Orlovskaya 2004a and b). The calibrated radiocarbon dates suggest that the Maikop ‘culture’ seems to have had a formative influence on steppe kurgan burial rituals and what now appears to be the later development of the Pit-Grave (Yamnaya) culture on the Eurasian steppes (Chernykh and Orlovskaya 2004a: 97).
...
In other words, sometime around the middle of the 4th millennium BCE or slightly subsequent to the initial appearance of the Maikop culture of the NW Caucasus, settlements containing proto-Kura-Araxes or early Kura-Araxes materials first appear across a broad area that stretches from the Caspian littoral of the northeastern Caucasus in the north to the Erzurum region of the Anatolian Plateau in the west. For simplicity’s sake these roughly simultaneous developments across this broad area will be considered as representing the beginnings of the Early Bronze Age or the initial stages of development of the KuraAraxes/Early Transcaucasian culture.
...
The ‘homeland’ (itself a very problematic concept) of the Kura-Araxes culture-historical community is difficult to pinpoint precisely, a fact that may suggest that there is no single well-demarcated area of origin, but multiple interacting areas including northeastern Anatolia as far as the Erzurum area, the catchment area drained by the Upper Middle Kura and Araxes Rivers in Transcaucasia and the Caspian corridor and adjacent mountainous regions of northeastern Azerbaijan and southeastern Daghestan. While broadly (and somewhat imprecisely) defined, these regions constitute on present evidence the original core area out of which the Kura-Araxes ‘culture-historical community’ emerged.
Kura-Araxes materials found in other areas are primarily intrusive in the local sequences. Indeed, many, but not all, sites in the Malatya area along the Upper Euphrates drainage of eastern Anatolia (e.g., Norsun-tepe, Arslantepe) and western Iran (e.g., Yanik Tepe, Godin Tepe) exhibit— albeit with some overlap—a relatively sharp break in material remains, including new forms of architecture and domestic dwellings, and such changes support the interpretation of a subsequent spread or dispersal from this broadly defined core area in the north to the southwest and southeast. The archaeological record seems to document a movement of peoples north to south across a very extensive part of the Ancient Near East from the end of the 4th to the first half of the 3rd millennium BCE. Although migrations are notoriously difficult to document on archaeological evidence, these materials constitute one of the best examples of prehistoric movements of peoples available for the Early Bronze Age.
Tel Aviv vol. 36, 2009 241–265
Origins, Homelands and Migrations: Situating the Kura-Araxes Early Transcaucasian ‘Culture’ within the History of Bronze Age Eurasia
Philip L. Kohl
This paper summarizes current understanding of the emergence, nature and subsequent southwestern and southeastern spread of the early Transcaucasian (eTC) or Kura-Araxes ‘culture-historical community’ (Russian: obshchnost’) and then places this complex cultural phenomenon in the context of the larger early Bronze Age world of the Ancient Near east and the western eurasian steppes.
Link
Recurrent Y-chromosome mutations from X-to-Y gene conversion
Forensic Sci Int Genet. 2013 Jun 26. pii: S1872-4973(13)00121-X. doi: 10.1016/j.fsigen.2013.05.010. [Epub ahead of print]
Multiple recurrent mutations at four human Y-chromosomal single nucleotide polymorphism sites in a 37bp sequence tract on the ARSDP1 pseudogene.
Niederstätter H, Berger B, Erhart D, Willuweit S, Geppert M, Gassner C, Schennach H, Parson W, Roewer L.
Abstract
The male-specific region of the human Y chromosome (MSY) is passed down clonally from father to son and mutation is the single driving force for Y-chromosomal diversification. The geographical distribution of MSY variation is non-random. Therefore, Y-chromosomal single nucleotide polymorphisms (Y-SNPs) are of forensic interest, as they can be utilized, e.g. for deducing the bio-geographical origin of biological evidence. This extra information can complement short tandem repeat data in criminal investigations. For forensic applications, however, any targeted marker has to be unequivocally interpretable. Here, we report findings for 17 samples from a population study comprising specimens from ∼3700 men living in Tyrol (Austria), indicating apparent homoplasic mutations at four Y-SNP loci on haplogroup R-M412/L51/S167, R-U152/S28, and L-M20 Y chromosomes. The affected Y-SNPs P41, P37, L202, and L203 mapped to a 37bp region on Yq11.21. Observing in multiple phylogenetic contexts up to four homoplasic mutations within such a short sequence tract is unlikely to result from a series of independent parallel mutations. Hence, we rather propose X-to-Y gene conversion as a more likely scenario. Practical implications arising from markers exhibiting paralogues on the Y chromosome or sites with a high propensity to recurrent mutation for database searches are addressed.
Link
Multiple recurrent mutations at four human Y-chromosomal single nucleotide polymorphism sites in a 37bp sequence tract on the ARSDP1 pseudogene.
Niederstätter H, Berger B, Erhart D, Willuweit S, Geppert M, Gassner C, Schennach H, Parson W, Roewer L.
Abstract
The male-specific region of the human Y chromosome (MSY) is passed down clonally from father to son and mutation is the single driving force for Y-chromosomal diversification. The geographical distribution of MSY variation is non-random. Therefore, Y-chromosomal single nucleotide polymorphisms (Y-SNPs) are of forensic interest, as they can be utilized, e.g. for deducing the bio-geographical origin of biological evidence. This extra information can complement short tandem repeat data in criminal investigations. For forensic applications, however, any targeted marker has to be unequivocally interpretable. Here, we report findings for 17 samples from a population study comprising specimens from ∼3700 men living in Tyrol (Austria), indicating apparent homoplasic mutations at four Y-SNP loci on haplogroup R-M412/L51/S167, R-U152/S28, and L-M20 Y chromosomes. The affected Y-SNPs P41, P37, L202, and L203 mapped to a 37bp region on Yq11.21. Observing in multiple phylogenetic contexts up to four homoplasic mutations within such a short sequence tract is unlikely to result from a series of independent parallel mutations. Hence, we rather propose X-to-Y gene conversion as a more likely scenario. Practical implications arising from markers exhibiting paralogues on the Y chromosome or sites with a high propensity to recurrent mutation for database searches are addressed.
Link
July 01, 2013
Complete mtDNA from Filipino groups

Complete mtDNA genomes of Filipino ethnolinguistic groups: a melting pot of recent and ancient lineages in the Asia-Pacific region
Frederick Delfin et al.
The Philippines is a strategic point in the Asia-Pacific region for the study of human diversity, history and origins, as it is a cross-road for human migrations and consequently exhibits enormous ethnolinguistic diversity. Following on a previous in-depth study of Y-chromosome variation, here we provide new insights into the maternal genetic history of Filipino ethnolinguistic groups by surveying complete mitochondrial DNA (mtDNA) genomes from a total of 14 groups (11 groups in this study and 3 groups previously published) including previously published mtDNA hypervariable segment (HVS) data from Filipino regional center groups. Comparison of HVS data indicate genetic differences between ethnolinguistic and regional center groups. The complete mtDNA genomes of 14 ethnolinguistic groups reveal genetic aspects consistent with the Y-chromosome, namely: diversity and heterogeneity of groups, no support for a simple dichotomy between Negrito and non-Negrito groups, and different genetic affinities with Asia-Pacific groups that are both ancient and recent. Although some mtDNA haplogroups can be associated with the Austronesian expansion, there are others that associate with South Asia, Near Oceania and Australia that are consistent with a southern migration route for ethnolinguistic group ancestors into the Asia-Pacific, with a timeline that overlaps with the initial colonization of the Asia-Pacific region, the initial colonization of the Philippines and a possible separate post-colonization migration into the Philippine archipelago.
Link